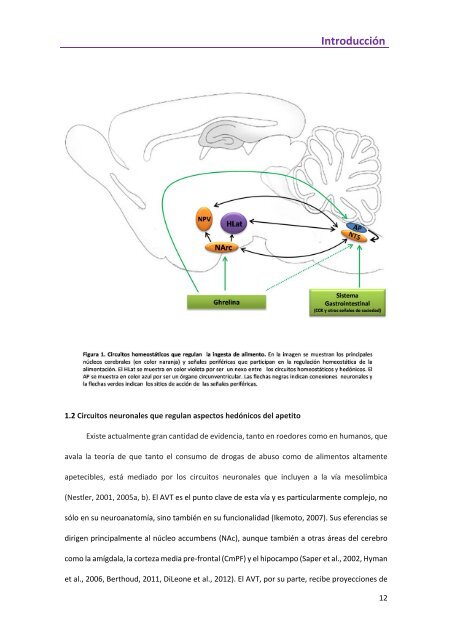
Introducción 1.2 Circuitos neuronales que regulan aspectos hedónicos del apetito Existe actualmente gran cantidad de evidencia, tanto en roedores como en humanos, que avala la teoría de que tanto el consumo de drogas de abuso como de alimentos altamente apetecibles, está mediado por los circuitos neuronales que incluyen a la vía mesolímbica (Nestler, 2001, 2005a, b). El AVT es el punto clave de esta vía y es particularmente complejo, no sólo en su neuroanatomía, sino también en su funcionalidad (Ikemoto, 2007). Sus eferencias se dirigen principalmente al núcleo accumbens (NAc), aunque también a otras áreas del cerebro como la amígdala, la corteza media pre‐frontal (CmPF) y el hipocampo (Saper et al., 2002, Hyman et al., 2006, Berthoud, 2011, DiLeone et al., 2012). El AVT, por su parte, recibe proyecciones de 12
Introducción varios núcleos, como el núcleo tegmental latero dorsal (LDTg) y el HLat, además de las áreas antes mencionadas que son inervadas por éste (Fadel and Deutch, 2002, Geisler and Zahm, 2005, Dickson et al., 2010). El neurotransmisor sintetizado predominantemente en el AVT es la dopamina, el cual se produce a partir del aminoácido L‐tirosina. El paso limitante de su síntesis es catalizado por la enzima tirosina hidroxilasa (TH), por lo que es utilizada como un marcador bioquímico para la identificación de las neuronas dopaminérgicas (Vucetic and Reyes, 2010). La dopamina interactúa con receptores de membrana pertenecientes a la familia de los receptores acoplados a proteína G, cuya activación lleva a la formación de segundos mensajeros y finalmente a la activación o represión de rutas de señalización específicas (Baik, 2013). Se conocen 5 subtipos de receptores dopaminérgicos los que, basados en sus propiedades estructurales y farmacológicas, se subdividen en 2 grupos: los de tipo D1, que estimulan los niveles intracelulares de AMP cíclico y comprenden los receptores D1 y D5 (Dearry et al., 1990, Zhou et al., 1990, Grandy et al., 1991, Sunahara et al., 1991) y los de tipo D2 que inhiben los niveles intracelulares de AMP cíclico y comprenden los receptores D2, D3 y D4 (Bunzow et al., 1988, Dal Toso et al., 1989, Sokoloff et al., 1990, Van Tol et al., 1991). Los receptores D1 y D2 son lo que se expresan más abundantemente en el cerebro (Baik, 2013). Los receptores D1 se encuentran exclusivamente en forma post‐sináptica en las neuronas inervadas por las neuronas dopaminérgicas, como las neuronas GABAérgicas del NAc. Los receptores D2 (y también los D3) se expresan no sólo a nivel post‐sináptico en células blanco de las neuronas dopaminérgicas sino también a nivel pre‐sináptico en neuronas dopaminérgicas (Sokoloff et al., 2006, Rondou et al., 2010, Beaulieu and Gainetdinov, 2011). En función de la distribución neuroanatómica de las neuronas dopaminérgicas, el AVT puede dividirse en al menos 3 subregiones: el núcleo paranigral (PN), el núcleo parabraquial pigmentado (PBP) y el núcleo interfascicular (IF), el cual es considerado por algunos autores como un núcleo independiente (Swanson, 1982, Ikemoto, 13
- Page 1 and 2: UNIVERSIDAD NACIONAL DE LA PLATA FA
- Page 3 and 4: Índice Abreviaturas……………
- Page 5 and 6: 5
- Page 7 and 8: Introducción específicas como el
- Page 9 and 10: Introducción Dentro de cada aspect
- Page 11: Introducción Las neuronas de los n
- Page 15 and 16: Introducción que eventualmente lle
- Page 17 and 18: Introducción datos sugieren que la
- Page 19 and 20: Introducción ACe, áreas corticale
- Page 21 and 22: Introducción La ghrelina es secret
- Page 23 and 24: Introducción Como se mencionó ant
- Page 25 and 26: Introducción muy frecuentes, es de
- Page 27 and 28: Introducción Por otro lado, se ha
- Page 29 and 30: Introducción 4. MODELOS EXPERIMENT
- Page 31 and 32: Introducción Hietala et al., 1994,
- Page 33 and 34: Objetivos OBJETIVO GENERAL El objet
- Page 35 and 36: 35
- Page 37 and 38: Materiales y Métodos la Comisión
- Page 39 and 40: Materiales y Métodos − − Tioni
- Page 41 and 42: Materiales y Métodos (Perello et a
- Page 43 and 44: Materiales y Métodos una cámara c
- Page 45 and 46: Materiales y Métodos inyectados bi
- Page 47 and 48: Materiales y Métodos derecha para
- Page 49 and 50: Materiales y Métodos 6.2.4 IHQ flu
- Page 51 and 52: Materiales y Métodos 6.3 Análisis
- Page 53 and 54: Materiales y Métodos corregidos po
- Page 55 and 56: Materiales y Métodos Para comparar
- Page 57 and 58: Resultados 1. ESTUDIO DE LOS CIRCUI
- Page 59 and 60: Resultados 59
- Page 61 and 62: Resultados 61
- Page 63 and 64:
Resultados En el AVT, también se o
- Page 65 and 66:
Resultados 1.3 Un único evento de
- Page 67 and 68:
Resultados el HLat fueron las que s
- Page 69 and 70:
Resultados 1.5 El bloqueo del recep
- Page 71 and 72:
Resultados Además de afectar la ca
- Page 73 and 74:
Resultados 1.6 Las neuronas product
- Page 75 and 76:
Resultados 75
- Page 77 and 78:
Resultados − Utilización de rato
- Page 79 and 80:
Resultados Con el objetivo de deter
- Page 81 and 82:
Resultados Por último, para determ
- Page 83 and 84:
Resultados 2. ESTUDIO DE LOS CIRCUI
- Page 85 and 86:
Resultados 2.2 La actividad locomot
- Page 87 and 88:
Resultados 87
- Page 89 and 90:
Resultados 89
- Page 91 and 92:
Resultados 91
- Page 93 and 94:
Resultados 93
- Page 95 and 96:
Resultados [caparazón medial: t(5)
- Page 97 and 98:
Resultados que para los animales de
- Page 99 and 100:
Resultados 2.9 La administración d
- Page 101 and 102:
Resultados 101
- Page 103 and 104:
Resultados 103
- Page 105 and 106:
105
- Page 107 and 108:
Discusión saciados, y que la expos
- Page 109 and 110:
Discusión mRNA de neuropéptidos y
- Page 111 and 112:
Discusión alimentación (Palmiter,
- Page 113 and 114:
Discusión neuronas productoras de
- Page 115 and 116:
Discusión 2004, Kelley et al., 200
- Page 117 and 118:
Discusión orexina no afectó el es
- Page 119 and 120:
Discusión ingirieron DRG en cuatro
- Page 121 and 122:
Discusión utilizadas, como el tipo
- Page 123 and 124:
Discusión Actualmente, existe evid
- Page 125 and 126:
Discusión La ingesta de DRG en for
- Page 127 and 128:
Discusión neuronas CRF de la amíg
- Page 129 and 130:
Discusión el sistema ghrelina‐GH
- Page 131 and 132:
Conclusiones En conclusión, los re
- Page 133 and 134:
133
- Page 135 and 136:
Bibliografía receptor‐mediated i
- Page 137 and 138:
Bibliografía Bugnon C, Fellmann D,
- Page 139 and 140:
Bibliografía Davis C, Levitan RD,
- Page 141 and 142:
Bibliografía Gama Sosa MA, De Gasp
- Page 143 and 144:
Bibliografía Hotta M, Ohwada R, Ak
- Page 145 and 146:
Bibliografía Kojima S, Nakahara T,
- Page 147 and 148:
Bibliografía Mason BL, Wang Q, Zig
- Page 149 and 150:
Bibliografía Palmiter RD (2007) Is
- Page 151 and 152:
Bibliografía Saper CB (1982) Conve
- Page 153 and 154:
Bibliografía Swart I, Jahng JW, Ov
- Page 155 and 156:
Bibliografía Zahm DS (2000) An int