

Smithsonian at the Poles: Contributions to International Polar
Smithsonian at the Poles: Contributions to International Polar
Smithsonian at the Poles: Contributions to International Polar

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
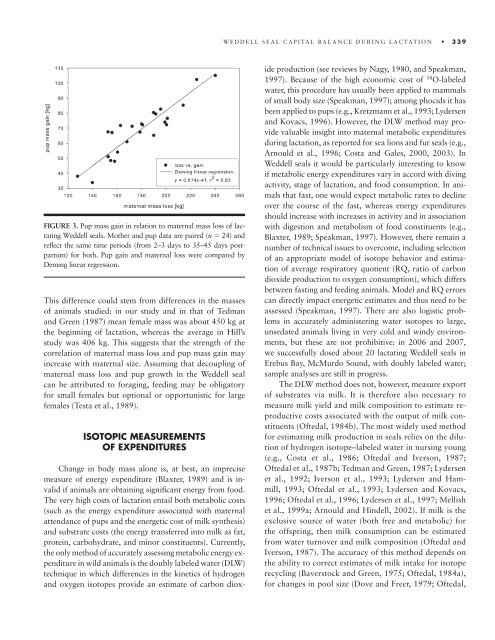
FIGURE 3. Pup mass gain in rel<strong>at</strong>ion <strong>to</strong> m<strong>at</strong>ernal mass loss of lact<strong>at</strong>ing<br />
Weddell seals. Mo<strong>the</strong>r and pup d<strong>at</strong>a are paired (n � 24) and<br />
refl ect <strong>the</strong> same time periods (from 2– 3 days <strong>to</strong> 35– 45 days postpartum)<br />
for both. Pup gain and m<strong>at</strong>ernal loss were compared by<br />
Deming linear regression.<br />
This difference could stem from differences in <strong>the</strong> masses<br />
of animals studied: in our study and in th<strong>at</strong> of Tedman<br />
and Green (1987) mean female mass was about 450 kg <strong>at</strong><br />
<strong>the</strong> beginning of lact<strong>at</strong>ion, whereas <strong>the</strong> average in Hill’s<br />
study was 406 kg. This suggests th<strong>at</strong> <strong>the</strong> strength of <strong>the</strong><br />
correl<strong>at</strong>ion of m<strong>at</strong>ernal mass loss and pup mass gain may<br />
increase with m<strong>at</strong>ernal size. Assuming th<strong>at</strong> decoupling of<br />
m<strong>at</strong>ernal mass loss and pup growth in <strong>the</strong> Weddell seal<br />
can be <strong>at</strong>tributed <strong>to</strong> foraging, feeding may be oblig<strong>at</strong>ory<br />
for small females but optional or opportunistic for large<br />
females (Testa et al., 1989).<br />
ISOTOPIC MEASUREMENTS<br />
OF EXPENDITURES<br />
Change in body mass alone is, <strong>at</strong> best, an imprecise<br />
measure of energy expenditure (Blaxter, 1989) and is invalid<br />
if animals are obtaining signifi cant energy from food.<br />
The very high costs of lact<strong>at</strong>ion entail both metabolic costs<br />
(such as <strong>the</strong> energy expenditure associ<strong>at</strong>ed with m<strong>at</strong>ernal<br />
<strong>at</strong>tendance of pups and <strong>the</strong> energetic cost of milk syn<strong>the</strong>sis)<br />
and substr<strong>at</strong>e costs (<strong>the</strong> energy transferred in<strong>to</strong> milk as f<strong>at</strong>,<br />
protein, carbohydr<strong>at</strong>e, and minor constituents). Currently,<br />
<strong>the</strong> only method of accur<strong>at</strong>ely assessing metabolic energy expenditure<br />
in wild animals is <strong>the</strong> doubly labeled w<strong>at</strong>er (DLW)<br />
technique in which differences in <strong>the</strong> kinetics of hydrogen<br />
and oxygen iso<strong>to</strong>pes provide an estim<strong>at</strong>e of carbon diox-<br />
WEDDELL SEAL CAPITAL BALANCE DURING LACTATION 339<br />
ide production (see reviews by Nagy, 1980, and Speakman,<br />
1997). Because of <strong>the</strong> high economic cost of 18 O-labeled<br />
w<strong>at</strong>er, this procedure has usually been applied <strong>to</strong> mammals<br />
of small body size (Speakman, 1997); among phocids it has<br />
been applied <strong>to</strong> pups (e.g., Kretzmann et al., 1993; Lydersen<br />
and Kovacs, 1996). However, <strong>the</strong> DLW method may provide<br />
valuable insight in<strong>to</strong> m<strong>at</strong>ernal metabolic expenditures<br />
during lact<strong>at</strong>ion, as reported for sea lions and fur seals (e.g.,<br />
Arnould et al., 1996; Costa and Gales, 2000, 2003). In<br />
Weddell seals it would be particularly interesting <strong>to</strong> know<br />
if metabolic energy expenditures vary in accord with diving<br />
activity, stage of lact<strong>at</strong>ion, and food consumption. In animals<br />
th<strong>at</strong> fast, one would expect metabolic r<strong>at</strong>es <strong>to</strong> decline<br />
over <strong>the</strong> course of <strong>the</strong> fast, whereas energy expenditures<br />
should increase with increases in activity and in associ<strong>at</strong>ion<br />
with digestion and metabolism of food constituents (e.g.,<br />
Blaxter, 1989; Speakman, 1997). However, <strong>the</strong>re remain a<br />
number of technical issues <strong>to</strong> overcome, including selection<br />
of an appropri<strong>at</strong>e model of iso<strong>to</strong>pe behavior and estim<strong>at</strong>ion<br />
of average respir<strong>at</strong>ory quotient (RQ, r<strong>at</strong>io of carbon<br />
dioxide production <strong>to</strong> oxygen consumption), which differs<br />
between fasting and feeding animals. Model and RQ errors<br />
can directly impact energetic estim<strong>at</strong>es and thus need <strong>to</strong> be<br />
assessed ( Speakman, 1997). There are also logistic problems<br />
in accur<strong>at</strong>ely administering w<strong>at</strong>er iso<strong>to</strong>pes <strong>to</strong> large,<br />
unsed<strong>at</strong>ed animals living in very cold and windy environments,<br />
but <strong>the</strong>se are not prohibitive: in 2006 and 2007,<br />
we successfully dosed about 20 lact<strong>at</strong>ing Weddell seals in<br />
Erebus Bay, McMurdo Sound, with doubly labeled w<strong>at</strong>er;<br />
sample analyses are still in progress.<br />
The DLW method does not, however, measure export<br />
of substr<strong>at</strong>es via milk. It is <strong>the</strong>refore also necessary <strong>to</strong><br />
measure milk yield and milk composition <strong>to</strong> estim<strong>at</strong>e reproductive<br />
costs associ<strong>at</strong>ed with <strong>the</strong> output of milk constituents<br />
(Oftedal, 1984b). The most widely used method<br />
for estim<strong>at</strong>ing milk production in seals relies on <strong>the</strong> dilution<br />
of hydrogen iso<strong>to</strong>pe– labeled w<strong>at</strong>er in nursing young<br />
(e.g., Costa et al., 1986; Oftedal and Iverson, 1987;<br />
Oftedal et al., 1987b; Tedman and Green, 1987; Lydersen<br />
et al., 1992; Iverson et al., 1993; Lydersen and Hammill,<br />
1993; Oftedal et al., 1993; Lydersen and Kovacs,<br />
1996; Oftedal et al., 1996; Lydersen et al., 1997; Mellish<br />
et al., 1999a; Arnould and Hindell, 2002). If milk is <strong>the</strong><br />
exclusive source of w<strong>at</strong>er (both free and metabolic) for<br />
<strong>the</strong> offspring, <strong>the</strong>n milk consumption can be estim<strong>at</strong>ed<br />
from w<strong>at</strong>er turnover and milk composition (Oftedal and<br />
Iverson, 1987). The accuracy of this method depends on<br />
<strong>the</strong> ability <strong>to</strong> correct estim<strong>at</strong>es of milk intake for iso<strong>to</strong>pe<br />
recycling (Bavers<strong>to</strong>ck and Green, 1975; Oftedal, 1984a),<br />
for changes in pool size (Dove and Freer, 1979; Oftedal,














