

Zwischen Naturschutz und Theoretischer Ökologie: Modelle zur ...
Zwischen Naturschutz und Theoretischer Ökologie: Modelle zur ...
Zwischen Naturschutz und Theoretischer Ökologie: Modelle zur ...

Erfolgreiche ePaper selbst erstellen
Machen Sie aus Ihren PDF Publikationen ein blätterbares Flipbook mit unserer einzigartigen Google optimierten e-Paper Software.
126 5 Dynamisches Multihabitatmodell – ein räumlich explizites Simulationsmodell<br />
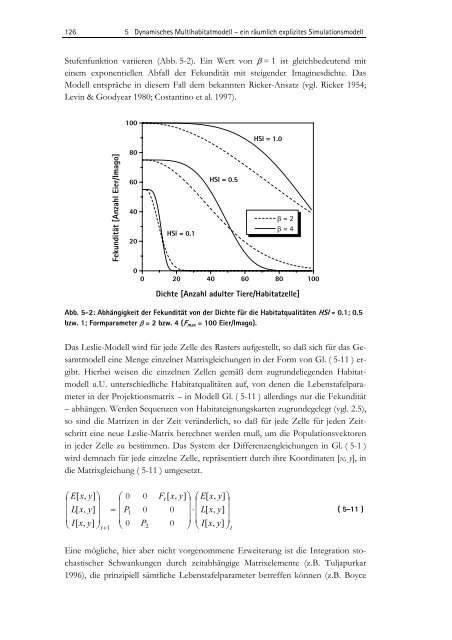
Stufenfunktion variieren (Abb. 5-2). Ein Wert von β = 1 ist gleichbedeutend mit<br />
einem exponentiellen Abfall der Fek<strong>und</strong>ität mit steigender Imaginesdichte. Das<br />
Modell entspräche in diesem Fall dem bekannten Ricker-Ansatz (vgl. Ricker 1954;<br />
Levin & Goodyear 1980; Costantino et al. 1997).<br />
Fek<strong>und</strong>ität [Anzahl Eier/Imago]<br />
100<br />
80<br />
60<br />
40<br />
20<br />
HSI = 0.1<br />
HSI = 0.5<br />
HSI = 1.0<br />
β = 2<br />
β = 4<br />
0<br />
0 20 40 60 80 100<br />
Dichte [Anzahl adulter Tiere/Habitatzelle]<br />
Abb. 5-2: Abhängigkeit der Fek<strong>und</strong>ität von der Dichte für die Habitatqualitäten HSI = 0.1; 0.5<br />
bzw. 1; Formparameter β = 2 bzw. 4 (Fmax = 100 Eier/Imago).<br />
Das Leslie-Modell wird für jede Zelle des Rasters aufgestellt, so daß sich für das Gesamtmodell<br />
eine Menge einzelner Matrixgleichungen in der Form von Gl. ( 5-11 ) ergibt.<br />
Hierbei weisen die einzelnen Zellen gemäß dem zugr<strong>und</strong>eliegenden Habitatmodell<br />
u.U. unterschiedliche Habitatqualitäten auf, von denen die Lebenstafelparameter<br />
in der Projektionsmatrix – in Modell Gl. ( 5-11 ) allerdings nur die Fek<strong>und</strong>ität<br />
– abhängen. Werden Sequenzen von Habitateignungskarten zugr<strong>und</strong>egelegt (vgl. 2.5),<br />
so sind die Matrizen in der Zeit veränderlich, so daß für jede Zelle für jeden Zeitschritt<br />
eine neue Leslie-Matrix berechnet werden muß, um die Populationsvektoren<br />
in jeder Zelle zu bestimmen. Das System der Differenzengleichungen in Gl. ( 5-1 )<br />
wird demnach für jede einzelne Zelle, repräsentiert durch ihre Koordinaten [x, y], in<br />
die Matrixgleichung ( 5-11 ) umgesetzt.<br />
⎛ E[<br />
x,<br />
y]<br />
⎞<br />
⎜ ⎟<br />
⎜ L[<br />
x,<br />
y]<br />
⎟<br />
⎜ I x y ⎟<br />
⎝ [ , ] ⎠<br />
t+<br />
1<br />
⎛ 0<br />
⎜<br />
= ⎜ P1<br />
⎜<br />
⎝ 0<br />
0<br />
0<br />
P<br />
2<br />
Ft<br />
[ x,<br />
y]<br />
⎞ ⎛ E[<br />
x,<br />
y]<br />
⎞<br />
⎟ ⎜ ⎟<br />
0 ⎟ ⋅ ⎜ L[<br />
x,<br />
y]<br />
⎟<br />
0 ⎟ ⎜ I x y ⎟<br />
⎠ ⎝ [ , ] ⎠<br />
t<br />
( 5-11 )<br />
Eine mögliche, hier aber nicht vorgenommene Erweiterung ist die Integration stochastischer<br />
Schwankungen durch zeitabhängige Matrixelemente (z.B. Tuljapurkar<br />
1996), die prinzipiell sämtliche Lebenstafelparameter betreffen können (z.B. Boyce



