

Biochemische Charakterisierung der siRNA-vermittelten Erkennung ...
Biochemische Charakterisierung der siRNA-vermittelten Erkennung ...
Biochemische Charakterisierung der siRNA-vermittelten Erkennung ...

Sie wollen auch ein ePaper? Erhöhen Sie die Reichweite Ihrer Titel.
YUMPU macht aus Druck-PDFs automatisch weboptimierte ePaper, die Google liebt.
elative Fluoreszenz<br />
5.7 <strong>Charakterisierung</strong> <strong>der</strong> <strong>siRNA</strong>-Bindung durch hAgo2<br />
Ratenkonstante betragen k 2 = 0,48 (± 0,23) s -1 und k 3 = 0,0282 (± 0,0184) s -1 (Mittelwerte aus<br />
fünf unabhängigen Experimenten). Die Ratenkonstanten <strong>der</strong> Assoziation von NHA-hAgo2 mit<br />
P-si2B-FAM sind demnach vergleichbar mit denen <strong>der</strong> Assoziation von NHA-hAgo2 und einer<br />
einzelsträngigen guide RNA.<br />
Im letzten Schritt wurde die Dissoziation des binären Komplexes aus NHA-hAgo2 und<br />
P-si2B-FAM unter pre-steady state Bedingungen untersucht. Dazu wurde P-si2B-FAM aus<br />
dem binären Komplex durch einen 100-fachen Überschuss nicht-markierter asLam verdrängt<br />
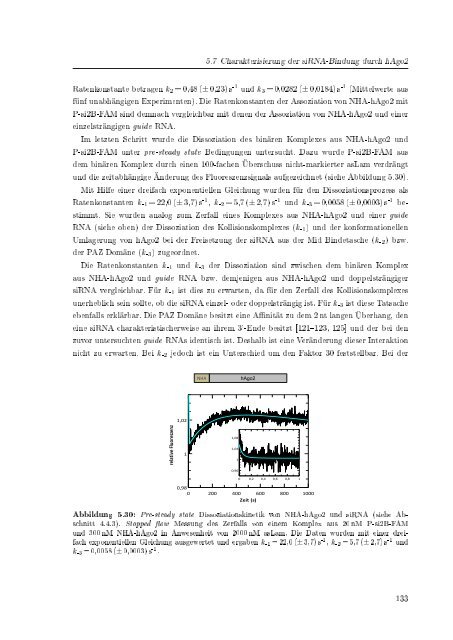
und die zeitabhängige Än<strong>der</strong>ung des Fluoreszenzsignals aufgezeichnet (siehe Abbildung 5.30).<br />
Mit Hilfe einer dreifach exponentiellen Gleichung wurden für den Dissoziationsprozess als<br />
Ratenkonstanten k -1 = 22,0 (± 3,7) s -1 , k -2 = 5,7 (± 2,7) s -1 und k -3 = 0,0058 (± 0,0003) s -1 bestimmt.<br />
Sie wurden analog zum Zerfall eines Komplexes aus NHA-hAgo2 und einer guide<br />
RNA (siehe oben) <strong>der</strong> Dissoziation des Kollisionskomplexes (k -1 ) und <strong>der</strong> konformationellen<br />
Umlagerung von hAgo2 bei <strong>der</strong> Freisetzung <strong>der</strong> <strong>siRNA</strong> aus <strong>der</strong> Mid Bindetasche (k -2 ) bzw.<br />
<strong>der</strong> PAZ Domäne (k -3 ) zugeordnet.<br />
Die Ratenkonstanten k -1 und k -3 <strong>der</strong> Dissoziation sind zwischen dem binären Komplex<br />
aus NHA-hAgo2 und guide RNA bzw. demjenigen aus NHA-hAgo2 und doppelsträngiger<br />
<strong>siRNA</strong> vergleichbar. Für k -1 ist dies zu erwarten, da für den Zerfall des Kollisionskomplexes<br />
unerheblich sein sollte, ob die <strong>siRNA</strong> einzel- o<strong>der</strong> doppelsträngig ist. Für k -3 ist diese Tatsache<br />
ebenfalls erklärbar. Die PAZ Domäne besitzt eine Anität zu dem 2 nt langen Überhang, den<br />
eine <strong>siRNA</strong> charakteristischerweise an ihrem 3'-Ende besitzt [121123, 125] und <strong>der</strong> bei den<br />
zuvor untersuchten guide RNAs identisch ist. Deshalb ist eine Verän<strong>der</strong>ung dieser Interaktion<br />
nicht zu erwarten. Bei k -2 jedoch ist ein Unterschied um den Faktor 30 feststellbar. Bei <strong>der</strong><br />
NHA<br />
hAgo2<br />
1,02<br />
1,08<br />
1<br />
1,04<br />
1<br />
0,96<br />
0 0,2 0,4 0,6 0,8 1<br />
0,98<br />
0 200 400 600 800 1000<br />
Zeit (s)<br />
Abbildung 5.30: Pre-steady state Dissoziationskinetik von NHA-hAgo2 und <strong>siRNA</strong> (siehe Abschnitt<br />
4.4.3). Stopped ow Messung des Zerfalls von einem Komplex aus 20 nM P-si2B-FAM<br />
und 300 nM NHA-hAgo2 in Anwesenheit von 2000 nM asLam. Die Daten wurden mit einer dreifach<br />
exponentiellen Gleichung ausgewertet und ergaben k -1 = 22,0 (± 3,7) s -1 , k -2 = 5,7 (± 2,7) s -1 und<br />
k -3 = 0,0058 (± 0,0003) s -1 .<br />
133













