

Principios de Neurociencia Haines 4a Ed_booksmedicos.org

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
414 Neurobiología <strong>de</strong> los sistemas<br />
Músculo loo circular<br />
Plexo mientónco<br />
Des<strong>de</strong> la<br />
Luz<br />
intestinal<br />
Motoneurona excitadora<br />
£ Contracción<br />
Intemeurona excitadora ascen<strong>de</strong>nte<br />
Neurona sensitiva<br />
Intemeurona excitadora <strong>de</strong>scen<strong>de</strong>nte<br />
Intemeurona inhibidora <strong>de</strong>scen<strong>de</strong>nte<br />
Motoneurona excitadora<br />
Distensión<br />
Retajaciór<br />
Motoneurona inhibidora<br />
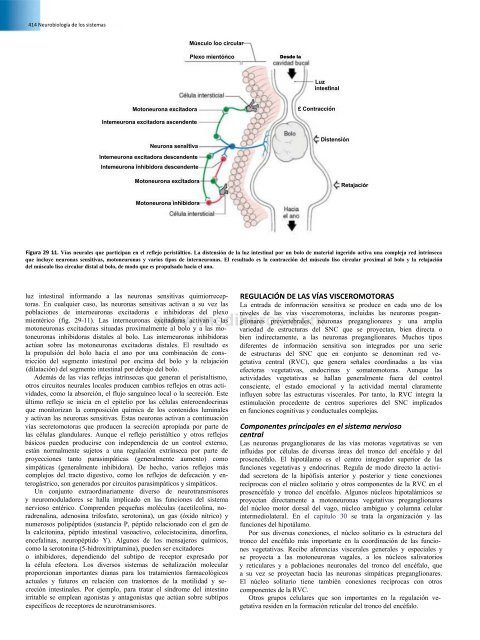
Figura 29 11. Vías neurales que participan en el reflejo peristáltico. La distensión <strong>de</strong> la luz intestinal por un bolo <strong>de</strong> material ingerido activa una compleja red intrínseca<br />
que incluye neuronas sensitivas, motoneuronas y varios tipos <strong>de</strong> interneuronas. El resultado es la contracción <strong>de</strong>l músculo liso circular proximal al bolo y la relajación<br />
<strong>de</strong>l músculo liso circular distal al bolo, <strong>de</strong> modo que es propulsado hacia el ano.<br />
luz intestinal informando a las neuronas sensitivas quimiorreceptoras.<br />
En cualquier caso, las neuronas sensitivas activan a su vez las<br />
poblaciones <strong>de</strong> interneuronas excitadoras e inhibidoras <strong>de</strong>l plexo<br />
mientérico (fig. 29-11). Las interneuronas excitadoras activan a las<br />
motoneuronas excitadoras situadas proximalmente al bolo y a las motoneuronas<br />
inhibidoras distales al bolo. Las interneuronas inhibidoras<br />
actúan sobre las motoneuronas excitadoras distales. El resultado es<br />
la propulsión <strong>de</strong>l bolo hacia el ano por una combinación <strong>de</strong> constricción<br />
<strong>de</strong>l segmento intestinal por encima <strong>de</strong>l bolo y la relajación<br />
(dilatación) <strong>de</strong>l segmento intestinal por <strong>de</strong>bajo <strong>de</strong>l bolo.<br />
A<strong>de</strong>más <strong>de</strong> las vías reflejas intrínsecas que generan el peristaltismo,<br />
otros circuitos neurales locales producen cambios reflejos en otras activida<strong>de</strong>s,<br />
como la absorción, el flujo sanguíneo local o la secreción. Este<br />
último reflejo se inicia en el epitelio por las células enteroendocrinas<br />
que monitorizan la composición química <strong>de</strong> los contenidos luminales<br />
y activan las neuronas sensitivas. Estas neuronas activan a continuación<br />
vías secretomotoras que producen la secreción apropiada por parte <strong>de</strong><br />
las células glandulares. Aunque el reflejo peristáltico y otros reflejos<br />
básicos pue<strong>de</strong>n producirse con in<strong>de</strong>pen<strong>de</strong>ncia <strong>de</strong> un control externo,<br />
están normalmente sujetos a una regulación extrínseca por parte <strong>de</strong><br />
proyecciones tanto parasimpáticas (generalmente aumento) como<br />
simpáticas (generalmente inhibidora). De hecho, varios reflejos más<br />
complejos <strong>de</strong>l tracto digestivo, como los reflejos <strong>de</strong> <strong>de</strong>fecación y enterogástrico,<br />
son generados por circuitos parasimpáticos y simpáticos.<br />
Un conjunto extraordinariamente diverso <strong>de</strong> neurotransmisores<br />
y neuromoduladores se halla implicado en las funciones <strong>de</strong>l sistema<br />
nervioso entérico. Compren<strong>de</strong>n pequeñas moléculas (acetilcolina, noradrenalina,<br />
a<strong>de</strong>nosina trifosfato, serotonina), un gas (óxido nítrico) y<br />
numerosos polipéptidos (sustancia P, péptido relacionado con el gen <strong>de</strong><br />
la calcitonina, péptido intestinal vasoactivo, colecistocinina, dinorfina,<br />
encefalinas, neuropéptido Y). Algunos <strong>de</strong> los mensajeros químicos,<br />
como la serotonina (5-hidroxitriptamina), pue<strong>de</strong>n ser excitadores<br />
o inhibidores, <strong>de</strong>pendiendo <strong>de</strong>l subtipo <strong>de</strong> receptor expresado por<br />
la célula efectora. Los diversos sistemas <strong>de</strong> señalización molecular<br />
proporcionan importantes dianas para los tratamientos farmacológicos<br />
actuales y futuros en relación con trastornos <strong>de</strong> la motilidad y secreción<br />
intestinales. Por ejemplo, para tratar el síndrome <strong>de</strong>l intestino<br />
irritable se emplean agonistas y antagonistas que actúan sobre subtipos<br />
específicos <strong>de</strong> receptores <strong>de</strong> neurotransmisores.<br />
REGULACIÓN DE LAS VÍAS VISCEROMOTORAS<br />
La entrada <strong>de</strong> información sensitiva se produce en cada uno <strong>de</strong> los<br />
niveles <strong>de</strong> las vías visceromotoras, incluidas las neuronas posganglionares<br />
prevertebrales, neuronas preganglionares y una amplia<br />
variedad <strong>de</strong> estructuras <strong>de</strong>l SNC que se proyectan, bien directa o<br />
bien indirectamente, a las neuronas preganglionares. Muchos tipos<br />
diferentes <strong>de</strong> información sensitiva son integrados por una serie<br />
<strong>de</strong> estructuras <strong>de</strong>l SNC que en conjunto se <strong>de</strong>nominan red vegetativa<br />
central (RVC), que genera señales coordinadas a las vías<br />
efectoras vegetativas, endocrinas y somatomotoras. Aunque las<br />
activida<strong>de</strong>s vegetativas se hallan generalmente fuera <strong>de</strong>l control<br />
consciente, el estado emocional y la actividad mental claramente<br />
influyen sobre las estructuras viscerales. Por tanto, la RVC integra la<br />
estimulación proce<strong>de</strong>nte <strong>de</strong> centros superiores <strong>de</strong>l SNC implicados<br />
en funciones cognitivas y conductuales complejas.<br />
Componentes principales en el sistema nervioso<br />
central<br />
Las neuronas preganglionares <strong>de</strong> las vías motoras vegetativas se ven<br />
influidas por células <strong>de</strong> diversas áreas <strong>de</strong>l tronco <strong>de</strong>l encéfalo y <strong>de</strong>l<br />
prosencéfalo. El hipotálamo es el centro integrador superior <strong>de</strong> las<br />
funciones vegetativas y endocrinas. Regula <strong>de</strong> modo directo la actividad<br />
secretora <strong>de</strong> la hipófisis anterior y posterior y tiene conexiones<br />
recíprocas con el núcleo solitario y otros componentes <strong>de</strong> la RVC en el<br />
prosencéfalo y tronco <strong>de</strong>l encéfalo. Algunos núcleos hipotalámicos se<br />
proyectan directamente a motoneuronas vegetativas preganglionares<br />
<strong>de</strong>l núcleo motor dorsal <strong>de</strong>l vago, núcleo ambiguo y columna celular<br />
intermediolateral. En el capítulo 30 se trata la <strong>org</strong>anización y las<br />
funciones <strong>de</strong>l hipotálamo.<br />
Por sus diversas conexiones, el núcleo solitario es la estructura <strong>de</strong>l<br />
tronco <strong>de</strong>l encéfalo más importante en la coordinación <strong>de</strong> las funciones<br />
vegetativas. Recibe aferencias viscerales generales y especiales y<br />
se proyecta a las motoneuronas vagales, a los núcleos salivatorios<br />
y reticulares y a poblaciones neuronales <strong>de</strong>l tronco <strong>de</strong>l encéfalo, que<br />
a su vez se proyectan hacia las neuronas simpáticas preganglionares.<br />
El núcleo solitario tiene también conexiones recíprocas con otros<br />
componentes <strong>de</strong> la RVC.<br />
Otros grupos celulares que son importantes en la regulación vegetativa<br />
resi<strong>de</strong>n en la formación reticular <strong>de</strong>l tronco <strong>de</strong>l encéfalo.
