- Page 2 and 3:
PRINCIPIOS DE Neurociencia Aplicaci
- Page 4 and 5:
PRINCIPIOS DE Neurociencia Aplicaci
- Page 6 and 7:
Colaboradores MARCH D. ARD, PhD Pro
- Page 8 and 9:
Prefacio La cuarta edición de Prin
- Page 10 and 11:
x Agradecimientos Gretchen, por cin
- Page 12 and 13: Sección CONCEPTOS BÁSICOS ~ d "'
- Page 14 and 15: Introducción a la estructura e ima
- Page 16 and 17: Introducción a la estructura e ima
- Page 18 and 19: Introducción a la estructura e ima
- Page 20 and 21: Conceptos básicos Figura 1-8. Imá
- Page 22 and 23: 10 Conceptos básicos estado y fuer
- Page 24 and 25: H 12 Conceptos básicos Figura 1-11
- Page 26 and 27: Introducción a la estructura e ima
- Page 28 and 29: Biología celular de las neuronas y
- Page 30 and 31: Biología celular de las neuronas y
- Page 32 and 33: Biología celular de las neuronas y
- Page 34 and 35: Biología celular de las neuronas y
- Page 36 and 37: Biología celular de las neuronas y
- Page 38 and 39: Biología celular de las neuronas y
- Page 40 and 41: Biología celular de las neuronas y
- Page 42 and 43: Biología celular de las neuronas y
- Page 44 and 45: Biología celular de las neuronas y
- Page 46 and 47: Capítulo 3 Base electroquímica de
- Page 48 and 49: 34 Conceptos básicos Tabla 3-2 Dis
- Page 50 and 51: 36 Conceptos básicos El potasio es
- Page 52 and 53: 38 Conceptos básicos C9 se agrega
- Page 54 and 55: 40 Conceptos básicos motora Mitoco
- Page 56 and 57: 42 Conceptos básicos En el caso de
- Page 58 and 59: 44 Conceptos básicos 9n» 0- Exter
- Page 60 and 61: 46 Conceptos básicos Figura 3-15.
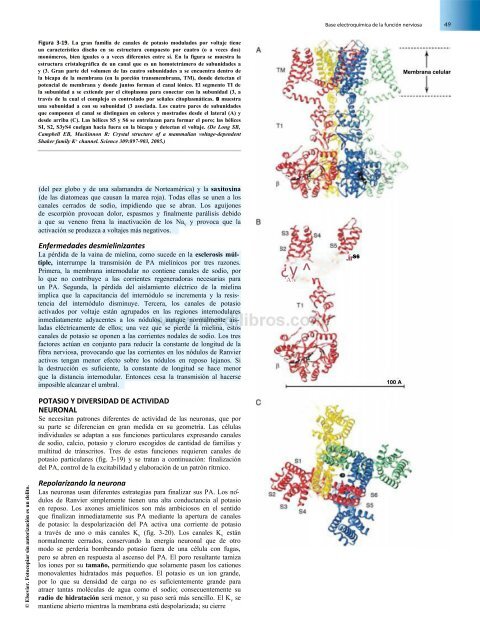
- Page 64 and 65: 50 Conceptos básicos está garanti
- Page 66 and 67: Base electroquímica de la función
- Page 68 and 69: Bases químicas de la comunicación
- Page 70 and 71: Bases químicas de la comunicación
- Page 72 and 73: Bases químicas de la comunicación
- Page 74 and 75: Bases químicas de la comunicación
- Page 76 and 77: Bases químicas de la comunicación
- Page 78 and 79: e-4 Conceptos básicos Bibliografí
- Page 80 and 81: 64 Conceptos básicos Placa neural
- Page 82 and 83: 66 Conceptos básicos Un encefaloce
- Page 84 and 85: 68 Conceptos básicos Flexura ponlm
- Page 86 and 87: 70 Conceptos básicos Telencélalo
- Page 88 and 89: 72 Conceptos básicos Dormatoma ♦
- Page 90 and 91: 74 Conceptos básicos C3 Modula esp
- Page 92 and 93: 76 Conceptos básicos Arteria coroi
- Page 94 and 95: 78 Conceptos básicos Figura 5-20.
- Page 96 and 97: 80 Conceptos básicos y es máxima
- Page 98 and 99: NEUROBIOLOG ÍA REGIONAL 6-16
- Page 100 and 101: Ventrículos, plexos coroideos y l
- Page 102 and 103: Ventrículos, plexos coroideos y l
- Page 104 and 105: Ventrículos, plexos coroideos y l
- Page 106 and 107: Ventrículos, plexos coroideos y l
- Page 108 and 109: Ventrículos, plexos coroideos y l
- Page 110 and 111: Ventrículos, plexos coroideos y l
- Page 112 and 113:
e-6 Neurobiología regional Bibliog
- Page 114 and 115:
96 Neurobiología regional Surco ne
- Page 116 and 117:
98 Neurobiología regional Seno sag
- Page 118 and 119:
100 Neurobiología regional Apófis
- Page 120 and 121:
102 Neurobiología regional el lado
- Page 122 and 123:
104 Neurobiología regional Hematom
- Page 124 and 125:
106 Neurobiología regional Cráneo
- Page 126 and 127:
Sangre entre tos lóbulos Cisterna
- Page 128 and 129:
Capítulo 8 Visión general del sis
- Page 130 and 131:
Visión general del sistema vascula
- Page 132 and 133:
Visión general del sistema vascula
- Page 134 and 135:
Visión general del sistema vascula
- Page 136 and 137:
Visión general del sistema vascula
- Page 138 and 139:
Visión general del sistema vascula
- Page 140 and 141:
Visión general del sistema vascula
- Page 142 and 143:
Visión general del sistema vascula
- Page 144 and 145:
Capítulo 9 Médula espinal D.E. Ha
- Page 146 and 147:
126 Neurobiología regional ESTRUCT
- Page 148 and 149:
128 Neurobiología regional Los cor
- Page 150 and 151:
130 Neurobiología regional Tracto
- Page 152 and 153:
132 Neurobiología regional Figura
- Page 154 and 155:
134 Neurobiología regional mononeu
- Page 156 and 157:
136 Neurobiología regional Tabla 9
- Page 158 and 159:
Médula espinal e-9 Bibliografía e
- Page 160 and 161:
Introducción al tronco del encéfa
- Page 162 and 163:
Introducción al tronco del encéfa
- Page 164 and 165:
Introducción al tronco del encéfa
- Page 166 and 167:
Introducción al tronco del encéfa
- Page 168 and 169:
Bulbo raquídeo 147 constituido por
- Page 170 and 171:
Bulbo raquídeo 149 I Fascículo gr
- Page 172 and 173:
Bulbo raquídeo 151 El núcleo reti
- Page 174 and 175:
Núcleo vestibular medial — Núcl
- Page 176 and 177:
Bulbo raquídeo 155 Arteria espinal
- Page 178 and 179:
Bulbo raquídeo 157 Dirección de l
- Page 180 and 181:
Puente y cerebelo 159 y de los núc
- Page 182 and 183:
Puente y cerebelo 161 Velo medular
- Page 184 and 185:
Puente y cerebelo 163 © Elsevier.
- Page 186 and 187:
Puente y cerebelo 165 Rodilla inter
- Page 188 and 189:
Fascículo longitudinal Sistema lec
- Page 190 and 191:
Capítulo 13 Mesencéfalo G.A. Mi h
- Page 192 and 193:
Mesencéfalo 171 Cuerpo geniculado
- Page 194 and 195:
Núcleo del troclear Fascículo lon
- Page 196 and 197:
Mesencéfalo 175 Decusación tegmen
- Page 198 and 199:
Mesencéfalo 177 en la unión entre
- Page 200 and 201:
Mesencéfalo 179 por efecto de la i
- Page 202 and 203:
e-10 Neurobiología regional Biblio
- Page 204 and 205:
182 Neurobiología regional forman
- Page 206 and 207:
184 Neurobiología regional vago (X
- Page 208 and 209:
186 Neurobiología regional La rela
- Page 210 and 211:
188 Neurobiología regional ñas EV
- Page 212 and 213:
190 Neurobiología regional ill Nú
- Page 214 and 215:
192 Neurobiología regional muscula
- Page 216 and 217:
I tienen sus somas en el ganglio de
- Page 218 and 219:
I cruzada carece de relevancia clí
- Page 220 and 221:
Resumen de los nervios craneales de
- Page 222 and 223:
Diencéfalo 199 Mesencéfalo Telenc
- Page 224 and 225:
Diencéfalo 201 . Quiasma Sustancia
- Page 226 and 227:
Diencefalo 203 Columna del fórnix
- Page 228 and 229:
Diencéfalo 205 Lóbulo ironu Giro
- Page 230 and 231:
Foramen interventricular Núdoo par
- Page 232 and 233:
arteria comunicante posterior y del
- Page 234 and 235:
e-12 Neurobiologia regional Bibliog
- Page 236 and 237:
212 Neurobiología regional Vesícu
- Page 238 and 239:
214 Neurobiología regional Giro pa
- Page 240 and 241:
216 Neurobiología regional Giros -
- Page 242 and 243:
218 Neurobiología regional Arteria
- Page 244 and 245:
220 Neurobiología regional fibras
- Page 246 and 247:
222 Neurobiología regional Rodilla
- Page 248 and 249:
224 Neurobiología regional Corona
- Page 250 and 251:
NEUROBIOLOGÍA DE LOS SISTEMAS 17 3
- Page 252 and 253:
Sistema somatosensitivo I: tacto di
- Page 254 and 255:
Sistema somatosensitivo I: tacto di
- Page 256 and 257:
Sistema somatosensitivo I: tacto di
- Page 258 and 259:
Sistema somatosensitivo I: tacto di
- Page 260 and 261:
Sistema somatosensitivo I: tacto di
- Page 262 and 263:
Sistema somatosensitivo I: tacto di
- Page 264 and 265:
Sistema somatosensitivo I: tacto di
- Page 266 and 267:
e-14 Neurobiología de los sistemas
- Page 268 and 269:
242 Neurobiología de los sistemas
- Page 270 and 271:
244 Neurobiología de los sistemas
- Page 272 and 273:
246 Neurobiología de los sistemas
- Page 274 and 275:
248 Neurobiología de los sistemas
- Page 276 and 277:
250 Neurobiología de los sistemas
- Page 278 and 279:
252 Neurobiología de los sistemas
- Page 280 and 281:
254 Neurobiología de los sistemas
- Page 282 and 283:
256 Neurobiología de los sistemas
- Page 284 and 285:
258 Neurobiología de los sistemas
- Page 286 and 287:
Sistema somatosensitivo II: nocicep
- Page 288 and 289:
VÍA ASCENDENTE DE LAS AFERENCIAS S
- Page 290 and 291:
Diversas áreas del córtex cerebra
- Page 292 and 293:
Vías sensitivas viscerales 265 Seg
- Page 294 and 295:
e-16 Neurobiología de los sistemas
- Page 296 and 297:
268 Neurobiología de los sistemas
- Page 298 and 299:
270 Neurobiología de los sistemas
- Page 300 and 301:
272 Neurobiología de los sistemas
- Page 302 and 303:
274 Neurobiología de los sistemas
- Page 304 and 305:
276 Neurobiología de los sistemas
- Page 306 and 307:
278 Neurobiología de los sistemas
- Page 308 and 309:
280 Neurobiología de los sistemas
- Page 310 and 311:
282 Neurobiología de los sistemas
- Page 312 and 313:
284 Neurobiología de los sistemas
- Page 314 and 315:
286 Neurobiología de los sistemas
- Page 316 and 317:
Capítulo 21 Sistema auditivo C.K.
- Page 318 and 319:
Pabellón auricular Estribo en la v
- Page 320 and 321:
Sistema auditivo 291 © Elsevier. F
- Page 322 and 323:
Sistema auditivo 293 Latencia (ms)
- Page 324 and 325:
Sistema auditivo 295 Figura 21-10.
- Page 326 and 327:
Sistema auditivo 297 indirectas pro
- Page 328 and 329:
de Brodmann. córtex auditivo |—C
- Page 330 and 331:
Capítulo 22 Sistema vestibular J.D
- Page 332 and 333:
302 Neurobiología de los sistemas
- Page 334 and 335:
304 Neurobiología de los sistemas
- Page 336 and 337:
306 Neurobiología de los sistemas
- Page 338 and 339:
308 Neurobiología de los sistemas
- Page 340 and 341:
310 Neurobiología de los sistemas
- Page 342 and 343:
312 Neurobiología de los sistemas
- Page 344 and 345:
Capítulo 23 Olfato y gusto K.L. Si
- Page 346 and 347:
Olfato y gusto 315 clulbo olfatorio
- Page 348 and 349:
Olfato y gusto 317 Giros orbitarios
- Page 350 and 351:
Olfato y gusto 319 © Elsevier. Fot
- Page 352 and 353:
Olfato y gusto 321 Faringe Epigkms
- Page 354 and 355:
Olfato y gusto 323 capaua ntei ía
- Page 356 and 357:
Capítulo 24 Sistema motor I: influ
- Page 358 and 359:
326 Neurobiología de los sistemas
- Page 360 and 361:
328 Neurobiología de los sistemas
- Page 362 and 363:
330 Neurobiología de los sistemas
- Page 364 and 365:
332 Neurobiología de los sistemas
- Page 366 and 367:
334 Neurobiología de los sistemas
- Page 368 and 369:
336 Neurobiología de los sistemas
- Page 370 and 371:
Sistema motor I: influencia sensiti
- Page 372 and 373:
Sistema motor II: sistemas corticó
- Page 374 and 375:
Sistema motor II: sistemas corticó
- Page 376 and 377:
Sistema motor II: sistemas corticó
- Page 378 and 379:
Sistema motor II: sistemas corticó
- Page 380 and 381:
Sistema motor II: sistemas corticó
- Page 382 and 383:
Sistema motor II: sistemas corticó
- Page 384 and 385:
Sistema motor II: sistemas corticó
- Page 386 and 387:
Sistema motor II: sistemas corticó
- Page 388 and 389:
Capítulo 26 Núcleos basales T.P.
- Page 390 and 391:
j 356 Neurobiología de los sistema
- Page 392 and 393:
358 Neurobiología de los sistemas
- Page 394 and 395:
360 Neurobiología de los sistemas
- Page 396 and 397:
362 Neurobiología de los sistemas
- Page 398 and 399:
364 Neurobiología de los sistemas
- Page 400 and 401:
366 Neurobiología de los sistemas
- Page 402 and 403:
368 Neurobiología de los sistemas
- Page 404 and 405:
Núcleos basales e-23 Bibliografía
- Page 406 and 407:
Tracto óptico Coliculo infenor Dec
- Page 408 and 409:
Cerebelo 373 Vormis Ungula Fisura p
- Page 410 and 411:
Cerebelo 375 (Mes) Lóbulo anterior
- Page 412 and 413:
Cerebelo 37 | = célula en cesta (C
- Page 414 and 415:
Cerebelo 379 ejemplo, se pueden reg
- Page 416 and 417:
Las singulares propiedades estructu
- Page 418 and 419:
Cerebelo 38: Espinocerebelo-zona de
- Page 420 and 421:
Pontocerebelo Córtex rr>otor Fibra
- Page 422 and 423:
Cerebelo 387 © Elsevier. Fotocopia
- Page 424 and 425:
e-24 Neurobiología de los sistemas
- Page 426 and 427:
390 Neurobiología de los sistemas
- Page 428 and 429:
392 Neurobiología de los sistemas
- Page 430 and 431:
394 Neurobiología de los sistemas
- Page 432 and 433:
396 Neurobiología de los sistemas
- Page 434 and 435:
398 Neurobiología de los sistemas
- Page 436 and 437:
400 Neurobiología de los sistemas
- Page 438 and 439:
402 Neurobiología de los sistemas
- Page 440 and 441:
404 Neurobiología de los sistemas
- Page 442 and 443:
Capítulo 29 Vías visceromotoras J
- Page 444 and 445:
Vías visceromotoras 407 Tabla 29-2
- Page 446 and 447:
posterior Ganglio raquídeo Hacia e
- Page 448 and 449:
Vías visceromotoras 411 Ganglio ra
- Page 450 and 451:
Vías visceromotoras 413 Codificaci
- Page 452 and 453:
Vías visceromotoras 415 © Elsevie
- Page 454 and 455:
e-26 Neurobiología de los sistemas
- Page 456 and 457:
418 Neurobiología de los sistemas
- Page 458 and 459:
420 Neurobiología de los sistemas
- Page 460 and 461:
422 Neurobiología de los sistemas
- Page 462 and 463:
424 Neurobiología de los sistemas
- Page 464 and 465:
426 Neurobiología de los sistemas
- Page 466 and 467:
428 Neurobiología de los sistemas
- Page 468 and 469:
430 Neurobiología de los sistemas
- Page 470 and 471:
Capítulo 31 Sistema límbico M.A.
- Page 472 and 473:
Córtex orbctolrontal caudal estruc
- Page 474 and 475:
Sistema límbico 435 Tracto Plexo c
- Page 476 and 477:
Sistema límbico 437 Figura 31-7. I
- Page 478 and 479:
Sistema límbico 439 Fórnix Estrí
- Page 480 and 481:
Sistema límbico 441 en la región
- Page 482 and 483:
Capítulo 32 Córtex cerebral J.C.
- Page 484 and 485:
444 Neurobiología de los sistemas
- Page 486 and 487:
446 Neurobiología de los sistemas
- Page 488 and 489:
448 Neurobiología de los sistemas
- Page 490 and 491:
450 Neurobiología de los sistemas
- Page 492 and 493:
452 Neurobiología de los sistemas
- Page 494 and 495:
Córtex cerebral e-29 Bibliografía
- Page 496 and 497:
Exploración neurológica 455 ■
- Page 498 and 499:
Exploración neurológica 457 $I Fi
- Page 500 and 501:
Exploración neurológica 459 A Fig
- Page 502 and 503:
Exploración neurológica 461 Figur
- Page 504 and 505:
Exploración neurológica 463 Figur
- Page 506 and 507:
Exploración neurológica 465 Figur
- Page 508 and 509:
Exploración neurológica 467 Figur
- Page 510 and 511:
índice alfabético A Abducción, m
- Page 512 and 513:
índice alfabético 471 © Elsevier
- Page 514 and 515:
índice alfabético 473 I © Elsevi
- Page 516 and 517:
índice alfabético 475 I © Elsevi
- Page 518 and 519:
índice alfabético 477 © Elsevier
- Page 520 and 521:
índice alfabético 479 © Elsevier
- Page 522 and 523:
índice alfabético 481 © Elsevier
- Page 524 and 525:
índice alfabético 483 © Elsevier
- Page 526 and 527:
índice alfabético 485 © Elsevier
- Page 528 and 529:
índice alfabético 487 I © Elsevi
- Page 530 and 531:
índice alfabético 489 © Elsevier
- Page 532 and 533:
índice alfabético 491 © Elsevier