

Principios de Neurociencia Haines 4a Ed_booksmedicos.org

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
50 Conceptos básicos<br />
está garantizado, ya que la corriente K v hará volver al potencial <strong>de</strong> membrana<br />
hacia V K/ el potencial <strong>de</strong> equilibrio para el potasio, repolarizando<br />
la membrana y permitiendo que Ky se cierre. El PA es así finalizado y la<br />
membrana retorna también a su estado <strong>de</strong> alta resistencia.<br />
UnM<br />
<strong>de</strong> reposo<br />
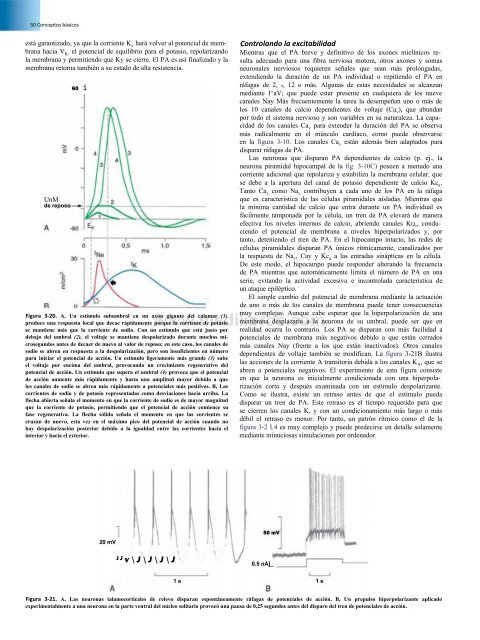
60 i<br />
Figura 3-20. A, Un estímulo subumbral en un axón gigante <strong>de</strong>l calamar (1)<br />
produce una respuesta local que <strong>de</strong>cae rápidamente porque la corriente <strong>de</strong> potasio<br />
se mantiene más que la corriente <strong>de</strong> sodio. Con un estímulo que está justo por<br />
<strong>de</strong>bajo <strong>de</strong>l umbral (2), el voltaje se mantiene <strong>de</strong>spolarizado durante muchos microsegundos<br />
antes <strong>de</strong> <strong>de</strong>caer <strong>de</strong> nuevo al valor <strong>de</strong> reposo; en este caso, los canales <strong>de</strong><br />
sodio se abren en respuesta a la <strong>de</strong>spolarización, pero son insuficientes en número<br />
para iniciar el potencial <strong>de</strong> acción. Un estímulo ligeramente más gran<strong>de</strong> (3) sube<br />
el voltaje por encima <strong>de</strong>l umbral, provocando un crecimiento regenerativo <strong>de</strong>l<br />
potencial <strong>de</strong> acción. Un estímulo que supera el umbral (4) provoca que el potencial<br />
<strong>de</strong> acción aumente más rápidamente y hasta una amplitud mayor <strong>de</strong>bido a que<br />
los canales <strong>de</strong> sodio se abren más rápidamente a potenciales más positivos. B, Las<br />
corrientes <strong>de</strong> sodio y <strong>de</strong> potasio representadas como <strong>de</strong>sviaciones hacia arriba. La<br />
flecha abierta señala el momento en que la corriente <strong>de</strong> sodio es <strong>de</strong> mayor magnitud<br />
que la corriente <strong>de</strong> potasio, permitiendo que el potencial <strong>de</strong> acción comience su<br />
fase regenerativa. La flecha sólida señala el momento en que las corrientes se<br />
cruzan <strong>de</strong> nuevo, esta vez en el máximo pico <strong>de</strong>l potencial <strong>de</strong> acción cuando no<br />
hay <strong>de</strong>spolarización posterior <strong>de</strong>bido a la igualdad entre las corrientes hacia el<br />
interior y hacia el exterior.<br />
Controlando la excitabilidad<br />
Mientras que el PA breve y <strong>de</strong>finitivo <strong>de</strong> los axones mielínicos resulta<br />
a<strong>de</strong>cuado para una fibra nerviosa motora, otros axones y somas<br />
neuronales nerviosos requieren señales que sean más prolongadas,<br />
extendiendo la duración <strong>de</strong> un PA individual o repitiendo el PA en<br />
ráfagas <strong>de</strong> 2, 6, 12 o más. Algunas <strong>de</strong> estas necesida<strong>de</strong>s se alcanzan<br />
mediante I^aV; que pue<strong>de</strong> estar presente en cualquiera <strong>de</strong> los nueve<br />
canales Nay Más frecuentemente la tarea la <strong>de</strong>sempeñan uno o más <strong>de</strong><br />
los 10 canales <strong>de</strong> calcio <strong>de</strong>pendientes <strong>de</strong> voltaje (Ca v ), que abundan<br />
por todo el sistema nervioso y son variables en su naturaleza. La capacidad<br />
<strong>de</strong> los canales Ca v para exten<strong>de</strong>r la duración <strong>de</strong>l PA se observa<br />
más radicalmente en el músculo cardíaco, como pue<strong>de</strong> observarse<br />
en la figura 3-10. Los canales Ca v están a<strong>de</strong>más bien adaptados para<br />
disparar ráfagas <strong>de</strong> PA.<br />
Las neuronas que disparan PA <strong>de</strong>pendientes <strong>de</strong> calcio (p. ej., la<br />
neurona piramidal hipocampal <strong>de</strong> la fig. 3-10C) poseen a menudo una<br />
corriente adicional que repolariza y estabiliza la membrana celular, que<br />
se <strong>de</strong>be a la apertura <strong>de</strong>l canal <strong>de</strong> potasio <strong>de</strong>pendiente <strong>de</strong> calcio Kc a .<br />
Tanto Ca v como Na v contribuyen a cada uno <strong>de</strong> los PA en la ráfaga<br />
que es característica <strong>de</strong> las células piramidales aisladas. Mientras que<br />
la mínima cantidad <strong>de</strong> calcio que entra durante un PA individual es<br />
fácilmente tamponada por la célula, un tren <strong>de</strong> PA elevará <strong>de</strong> manera<br />
efectiva los niveles internos <strong>de</strong> calcio, abriendo canales Kq,, conduciendo<br />
el potencial <strong>de</strong> membrana a niveles hiperpolarizados y, por<br />
tanto, <strong>de</strong>teniendo el tren <strong>de</strong> PA. En el hipocampo intacto, las re<strong>de</strong>s <strong>de</strong><br />
células piramidales disparan PA únicos rítmicamente, canalizados por<br />
la respuesta <strong>de</strong> Na v , Cay y Kc a a las entradas sinápticas en la célula.<br />
De este modo, el hipocampo pue<strong>de</strong> respon<strong>de</strong>r alterando la frecuencia<br />
<strong>de</strong> PA mientras que automáticamente limita el número <strong>de</strong> PA en una<br />
serie, evitando la actividad excesiva e incontrolada característica <strong>de</strong><br />
un ataque epiléptico.<br />
El simple cambio <strong>de</strong>l potencial <strong>de</strong> membrana mediante la actuación<br />
<strong>de</strong> uno o más <strong>de</strong> los canales <strong>de</strong> membrana pue<strong>de</strong> tener consecuencias<br />
muy complejas. Aunque cabe esperar que la hiperpolarización <strong>de</strong> una<br />
membrana <strong>de</strong>splazaría a la neurona <strong>de</strong> su umbral, pue<strong>de</strong> ser que en<br />
realidad ocurra lo contrario. Los PA se disparan con más facilidad a<br />
potenciales <strong>de</strong> membrana más negativos <strong>de</strong>bido a que están cerrados<br />
más canales Nay (frente a los que están inactivados). Otros canales<br />
<strong>de</strong>pendientes <strong>de</strong> voltaje también se modifican. La figura 3-21B ilustra<br />
las acciones <strong>de</strong> la corriente A transitoria <strong>de</strong>bida a los canales K A , que se<br />
abren a potenciales negativos. El experimento <strong>de</strong> esta figura consiste<br />
en que la neurona es inicialmente condicionada con una hiperpolarización<br />
corta y <strong>de</strong>spués examinada con un estímulo <strong>de</strong>spolarizante.<br />
Como se ilustra, existe un retraso antes <strong>de</strong> que el estímulo pueda<br />
disparar un tren <strong>de</strong> PA. Este retraso es el tiempo requerido para que<br />
se cierren los canales K, y con un condicionamiento más largo o más<br />
débil el retraso es menor. Por tanto, un patrón rítmico como el <strong>de</strong> la<br />
figura 3-2 L4 es muy complejo y pue<strong>de</strong> pre<strong>de</strong>cirse en <strong>de</strong>talle solamente<br />
mediante minuciosas simulaciones por or<strong>de</strong>nador.<br />
20 mV<br />
J J v\J\J\J\J<br />
50 mV<br />
0.5 nA]_<br />
1 s<br />
1 s<br />
Figura 3-21. A, Las neuronas talamocorticales <strong>de</strong> relevo disparan espontáneamente ráfagas <strong>de</strong> potenciales <strong>de</strong> acción. B, Un prepulso hiperpolarizante aplicado<br />
experimentalmente a una neurona en la parte ventral <strong>de</strong>l núcleo solitario provocó una pausa <strong>de</strong> 0,25 segundos antes <strong>de</strong>l disparo <strong>de</strong>l tren <strong>de</strong> potenciales <strong>de</strong> acción.
